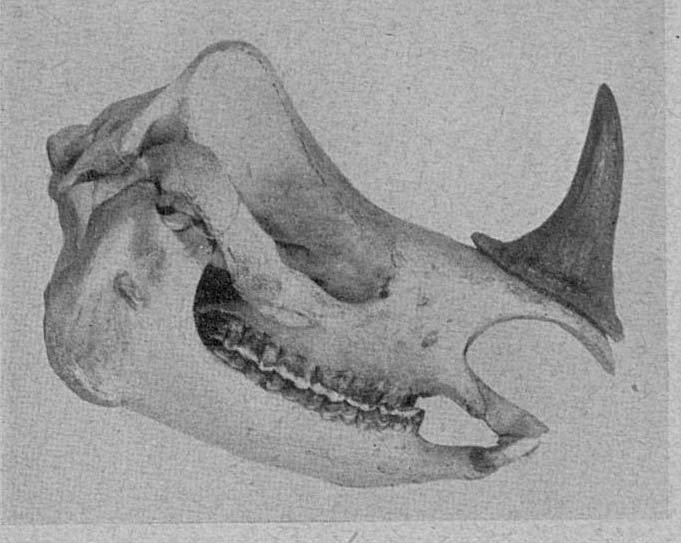
This hand colored steel engraving appeared in a children’s reader published by William Darton at the Repertory of Genius, 58 Holborn Hill, London

The world's largest online rhinoceros library dedicated to assisting research and conservation efforts globally.
 Image
ImageThis hand colored steel engraving appeared in a children’s reader published by William Darton at the Repertory of Genius, 58 Holborn Hill, London
 Related images
Related images